

Imágenes cedidas por los autores
El calentamiento global de origen antrópico provoca cambios en las dinámicas de los ecosistemas terrestres y acuáticos en todas las regiones climáticas de Europa. Una de las consecuencias de estas alteraciones es el incremento del riesgo de enfermedades transmitidas por vectores, que afectan tanto a los animales como a las poblaciones humanas. Los cambios en la hidrología y el aumento de la temperatura ambiental influyen directamente en los vectores, de manera que pueden aumentar tanto su rango de distribución como sus periodos de actividad a lo largo del año.
Epidemiología
Una de estas enfermedades es la dirofilariosis cardiopulmonar, una enfermedad zoonósica cuyo agente causal es el nematodo parásito Dirofilaria immitis.
Hospedadores
Los cánidos y los félidos, tanto domésticos como silvestres, son sus hospedadores definitivos.
Otros mamíferos, como mustélidos y carnívoros silvestres (lobo, zorros, lince ibérico, etc.) también pueden verse afectados en nuestro entorno.
En los humanos, considerados hospedadores accidentales, la infección suele ser benigna y asintomática, aunque ocasionalmente se manifiesta mediante la aparición de nódulos pulmonares calcificados.
Vectores
Los mosquitos culícidos de los géneros Culex, Aedes, Anopheles, Coquillettidia y Mansonia, entre otros, actúan como vectores.
En España y Portugal, Cx. pipiens y Cx. theileri son consideradas las especies más importantes en la transmisión de D. immitis, ya que son las más abundantes y ampliamente distribuidas en el territorio. Ambas han sido reportadas como transmisoras de D. immitis en la península ibérica, y solo Cx. theileri en las Islas Canarias y Madeira.
Ciclo biológico
Los vermes adultos de D. immitis se localizan entre la arteria pulmonar y el ventrículo derecho del corazón de su hospedador definitivo. Las hembras ovovivíparas liberan microfilarias al torrente sanguíneo.
Cuando un mosquito hembra se alimenta de sangre del hospedador infectado, ingiere estas microfilarias, que sufren dos mudas en su interior: de microfilaria a larva 2 (L2) y de L2 a L3, que son las larvas infectantes.
Este desarrollo larvario en el interior del vector, también denominado incubación extrínseca, es un proceso dependiente de temperatura:
- El umbral térmico mínimo para el desarrollo de las larvas es de 14ºC.
- Por debajo de esta temperatura, el crecimiento de las larvas se interrumpe, aunque pueden preservar su viabilidad durante algunos días.
Las L3 migran al aparato bucal del vector y son inoculadas en el hospedador definitivo durante la siguiente ingestión de sangre. Penetran rápidamente por la herida causada por la picadura del vector y entran en el torrente sanguíneo, donde mudan a L4 y L5 (adultos inmaduros). Estas llegan a la arteria pulmonar, donde alcanzan la madurez sexual completando el ciclo biológico hacia los 4-6 meses después de la infección.
Factores de riesgo y transmisión
Es una enfermedad cosmopolita especialmente sensible a los cambios en el clima. Variables como el cambio climático, el desarrollo urbano y periurbano (efecto isla de calor), la presencia de cultivos de regadío y la formación de masas de agua estancada generan microclimas óptimos para la cría de los mosquitos, que favorecen la idoneidad del hábitat para la enfermedad.
Además, el tránsito constante de mascotas y reservorios microfilarémicos entre áreas endémicas y zonas libres de la enfermedad, la falta de quimioprofilaxis y la presencia de carnívoros silvestres, que actúan como reservorios naturales, favorecen la transmisión.
Distribución y prevalencia
En Europa, durante los últimos años, se ha expandido desde los países tradicionalmente endémicos de la cuenca mediterránea y del sudeste del continente hasta zonas del centro-norte de Europa, donde actualmente se considera endémica en muchos países.
En España, la prevalencia media es del 6,47 %:
- Las zonas con mayor prevalencia son las Islas Canarias y las Islas Baleares —Tenerife (17,32 %), Ibiza (17,09 %), Gran Canaria (16,03 %), La Palma (15,65 %), La Gomera (11,54 %) y Mallorca (11,24 %) —seguidas de las provincias de Cádiz (13,68 %), Pontevedra (12,61 %) y Huelva (11,11 %).
- En el suroeste y en el centro-oeste y la costa mediterránea las prevalencias varían del 5 al 10 %.
- En las provincias del norte, centro-este y sudeste del país se observan prevalencias menores del 5 %.
- En Portugal, la prevalencia en perros es del 5,9 %:
- Los distritos con mayor prevalencia incluyen Aveiro (15,0 %), Coímbra (9,9 %), Beja (9,7 %) y Faro (8,8 %), seguidos de Setúbal (8,7 %), Castelo Branco (7,4 %), Viana do Castelo (6,3 %) y la isla de Madeira (5,8 %).
- Por el contrario, las prevalencias más bajas se encuentran en Vila Real (1,4 %), Porto (2,9 %), Guarda (3,1 %) y Braganza (3,3 %).
- Las islas Azores son la única región donde, hasta el momento, no se han encontrado perros positivos a D. immitis.
Mapas de riesgo de infección como herramienta de control
La distribución geográfica de la mayoría de las parasitosis transmitidas por vectores no es un fenómeno sujeto al azar, sino predecible en el espacio y en el tiempo. La transmisión del parásito depende estrechamente de los vectores, las condiciones climáticas y la presencia de hospedadores definitivos, imprescindibles para completar su ciclo biológico.
En la actualidad, para representar esta compleja epidemiología, se han adoptado herramientas ecoinformáticas destinadas a la creación de mapas de riesgo de infección colorimétricos, que identifican las zonas de mayor vulnerabilidad y las áreas donde la enfermedad podría establecerse.
Metodología y herramientas (MNE)
Los modelos de nicho ecológico (MNE) son algoritmos estadísticos que correlacionan los puntos de presencia georreferenciados de una especie con una serie de variables ambientales para estimar las condiciones en las que un ser vivo puede vivir y reproducirse.
Crear mapas de riesgo para la dirofilariosis cardiopulmonar a partir de esta metodología es un proceso complejo en el que se deben tener en cuenta dos aspectos: la idoneidad de hábitat del vector y los limitantes fisiológicos del parásito dentro de este (incubación extrínseca), tanto a escala anual como mensual.
Idoneidad de hábitat del vector
Para generar mapas colorimétricos de riesgo de infección para la dirofilariosis cardiopulmonar en España y Portugal, el primer paso es determinar la idoneidad de hábitat de los vectores Cx. pipiens y Cx. theileri a partir del algoritmo MaxEnt, con variables medioambientales de alta resolución (1 km2) relacionadas con la biología de los vectores.
Estas variables contienen información relacionada con la temperatura, precipitaciones, huella humana, ríos, masas de agua, cultivos de regadío, densidad de arbustos y densidad de herbáceas. Todas ellas se recortaron con una misma extensión y coordenadas a partir del software QGIS.
Limitantes fisiológicos del parásito
Para incluir un limitante fisiológico del parasito en el mapa de riesgo, se calcula el número de generaciones de D. immitis en el vector en función de la temperatura. Este método, descrito por Genchi et al. (2005), se adapta para calcular el número de generaciones por km2 en cada mes del año, además de la media anual.
Este método se basa en la acumulación de grados-día (Growing Degree Days, GDDs) durante los días en los que la temperatura media diaria supera los 14 °C, umbral mínimo necesario para el desarrollo de la larva L3 del parásito en el mosquito. Se considera que una generación completa se produce cuando se acumulan al menos 130 GDDs en un periodo máximo de 30 días (vida media del mosquito).
Dinámicas del riesgo
Península ibérica y Baleares
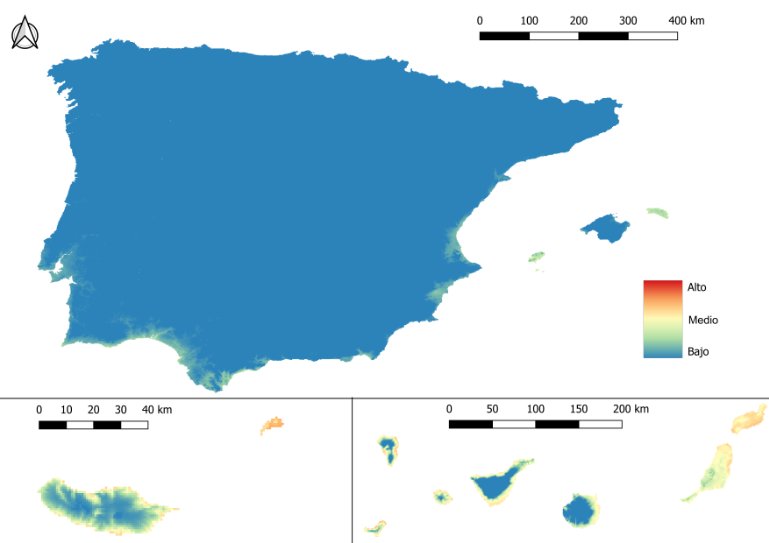
El mapa de riesgo anual de infección para D. immitis en la península ibérica y las Islas Baleares indica que el riesgo varía de moderado a alto en grandes extensiones del territorio donde el hábitat es favorable para los vectores y las temperaturas permiten un elevado número de generaciones del parásito (imagen 1):
- El mayor riesgo de infección se localiza a lo largo de la costa oriental, el sur peninsular y las Islas Baleares, así como en las áreas cercanas a las cuencas de los principales ríos.
- El riesgo es muy bajo en las zonas de mayor altitud, como los sistemas montañosos.
Respecto a la estacionalidad, el riesgo de infección sigue una dinámica muy marcada como consecuencia de las variaciones de temperatura, un factor imprescindible para el desarrollo del parásito y la actividad de los mosquitos vectores:
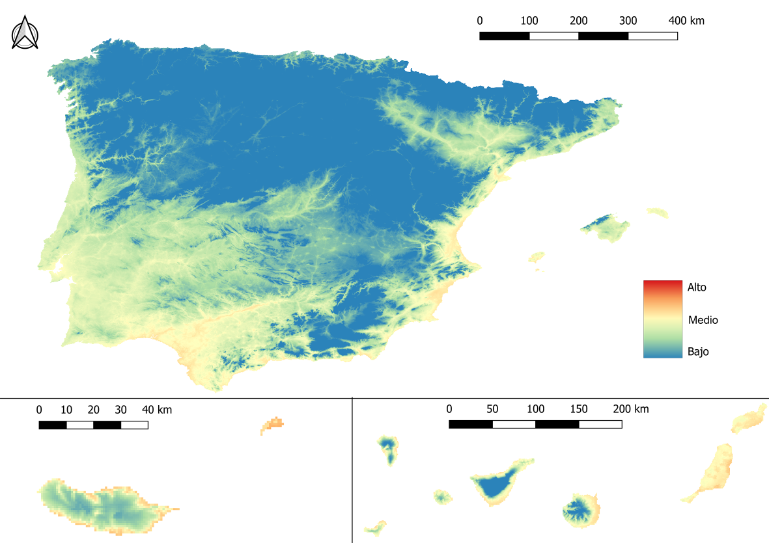
- De enero a marzo, el riesgo es muy bajo en prácticamente todo el territorio, aunque la transmisión no desaparece totalmente (imagen 2). En las zonas costeras del sur, el Algarve y focos específicos del litoral mediterráneo, se mantiene riesgo bajo/medio incluso durante el invierno debido a las temperaturas más suaves. En marzo, este riesgo se extiende por la cuenca del Guadalquivir cada vez más hacia el interior.
- Desde abril hasta junio, se observa una gran expansión desde las costas hacia el interior, de manera que se llega a riesgo medio/alto en las Islas Baleares y los valles del Guadalquivir, Guadiana y Ebro (imagen 3).
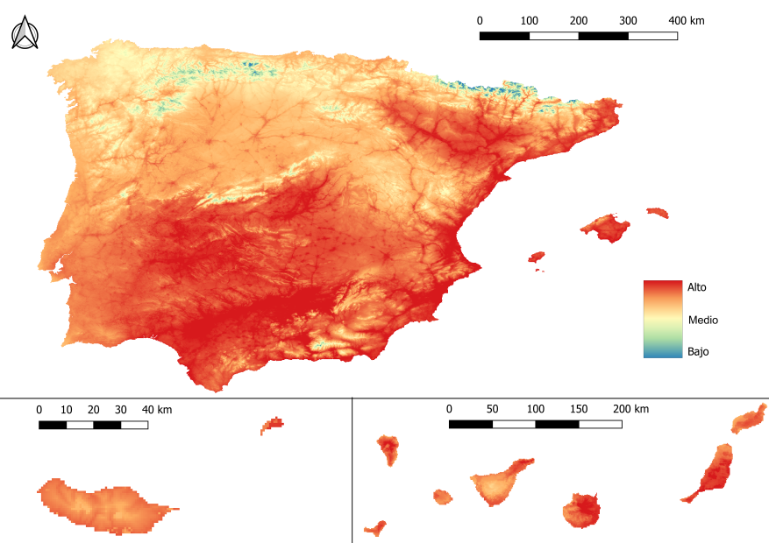
- El máximo riesgo se alcanza entre julio y septiembre, cuando gran parte de la mitad sur y las regiones costeras del levante se sitúan en niveles de riesgo muy alto (imagen 4). En la mitad norte de la península el riesgo llega a ser medio/alto a excepción de las cordilleras, donde el riesgo es menor como consecuencia de la altitud.
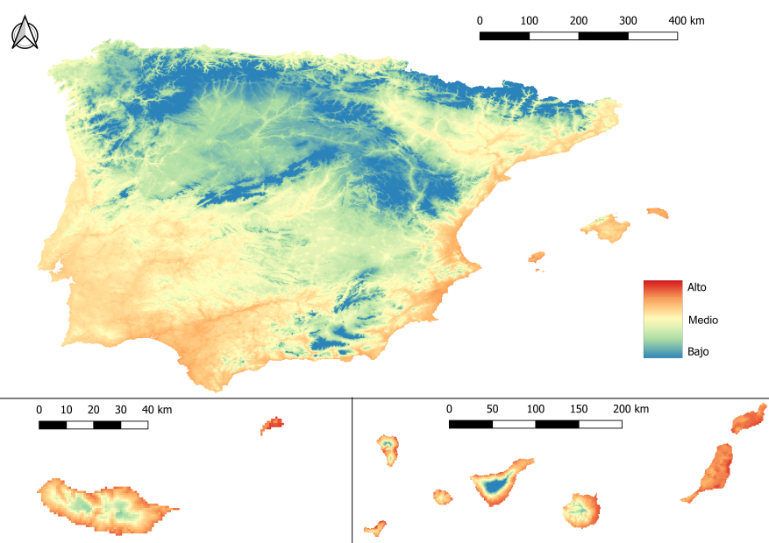
- Entre octubre y diciembre, el riesgo disminuye progresivamente desde la meseta central y las zonas de montaña hacia el litoral (imagen 5).
Islas Canarias
En las Islas Canarias el riesgo también alcanza su máximo valor en verano, aunque se mantiene moderado en la costa durante el invierno, lo que indica un riesgo de infección sostenido durante todo el año, condicionado por la altitud (imágenes 2-5):
- De enero a marzo, el riesgo es bajo en las medianías y las cumbres a excepción de la costa, donde se mantiene moderado.
- De abril a junio, se observa una expansión del riesgo hacia las medianías.
- El máximo riesgo se alcanza entre julio y septiembre, cuando la mayor parte del territorio se sitúa en riesgo muy alto.
- Entre octubre y diciembre, el riesgo retrocede gradualmente hacia el litoral.
A diferencia de zonas continentales como la península, la persistencia de niveles medios/altos en las costas durante todo el año confirma que el archipiélago mantiene una ventana de transmisión ininterrumpida en bajas altitudes.
Madeira
La dinámica mensual del riesgo de infección en Madeira es similar a la de las Islas Canarias debido a que presentan características comunes al tratarse de archipiélagos subtropicales (imágenes 2-5):
- Durante los meses de enero, febrero, marzo y abril el riesgo se mantiene muy bajo en las zonas de altitud y medianías, con algún foco de riesgo aislado en la costa.
- En mayo, mientras el riesgo del interior montañoso sigue siendo muy bajo, las zonas costeras aumentan a riesgo medio.
- A partir de junio se observa una progresión rápida hacia los máximos valores de riesgo con una tendencia ascendente. Durante los meses de julio y agosto el riesgo aumenta de forma drástica no solo en la costa, sino también en las medianías y zonas de mayor altitud.
- En el último cuatrimestre del año se produce un retroceso del riesgo debido a la vuelta del invierno. Durante septiembre aún se conservan niveles medios/altos de riesgo en las costas. En octubre continúa disminuyendo el riesgo desde el interior hacia el exterior, hasta alcanzar niveles muy bajos en invierno.
Azores
En el archipiélago de las Azores, el riesgo de infección por D. immitis presenta una dinámica marcadamente estacional, condicionado por su clima oceánico templado:
- Durante casi todo el primer semestre el riesgo es bajo en todo el archipiélago, ya que las temperaturas no alcanzan el umbral crítico para la incubación extrínseca del parásito.
- El riesgo comienza a aumentar en las zonas de menor altitud en junio, y los valores de riesgo en medio/alto se alcanzan en julio y agosto.
- A partir de septiembre el riesgo comienza a descender hasta alcanzar mínimos en el último trimestre del año.
En comparación con otros archipiélagos subtropicales, como Canarias y Madeira, la ventana de transmisión en Azores es más estrecha debido a su clima menos favorable, consecuencia de su localización en el Atlántico norte.
Validación del modelo y aplicaciones
Para determinar la capacidad predictiva de los mapas de riesgo, es necesario validarlos a partir de datos reales de la enfermedad. Para ello se geolocalizaron perros infectados y se superpusieron sobre el mapa de riesgo medio anual, categorizado en cinco clases de riesgo (“muy alto”, “alto”, “medio”, “bajo” y “muy bajo”):
- En la península ibérica, se observó que los perros infectados estaban geolocalizados en zonas de riesgo muy alto o alto (73 %), distribuidos a lo largo de las costas mediterránea y atlántica meridional y extendiéndose hacia el interior a través de las cuencas de los principales sistemas fluviales. Por el contrario, las regiones de la meseta central presentaban predominantemente un riesgo medio (26 %) y las de gran altitud, un riesgo bajo.
- Tanto las Islas Canarias como las Islas Baleares presentaban una elevada densidad de casos positivos, en consonancia con los altos valores de riesgo observados. En las Islas Baleares, todos los animales infectados estaban localizados en zonas de muy alto o alto riesgo. En las Islas Canarias el mayor número de animales (75 %) se encontraba en zonas costeras (zonas de medianía) y el resto, en zonas centrales de mayor y media altitud, con riesgo medio/bajo.
- Madeira presentaba un patrón de riesgo alto, concentrado principalmente a lo largo de su perímetro costero, y no se registraron casos en zonas de riesgo medio, bajo o muy bajo.
- En Azores, por el momento ausente de la enfermedad, no se geolocalizó ningún punto positivo.
Gracias a esta validación podemos confirmar la fiabilidad de la herramienta para predecir la realidad epidemiológica en todos los territorios analizados.
Conclusión
Estos mapas confirman que el riesgo de infección por D. immitis es constante en España y Portugal.
Sin embargo, no es uniforme: las zonas con mayor riesgo son las costas del sur y el este peninsular, así como la mayor parte de los archipiélagos, mientras que la altitud actúa como un protector natural disminuyendo el riesgo. Además, este riesgo aumenta durante los meses de verano, especialmente julio y agosto, mientras que disminuye durante los meses de invierno, por lo que las medidas para controlar la infección deben implementarse según la época del año.
Esta herramienta está diseñada para que cuidadores de mascotas y profesionales veterinarios tengan una guía clara sobre cuándo reforzar la prevención frente a la enfermedad. Bajo el enfoque One Health, el modelo destaca la importancia de la dirofilariosis no solo para los animales, sino como un desafío de salud pública. Esta metodología permite establecer una herramienta sólida, de alta resolución espacial, con el fin de implementar medidas reales de control de la enfermedad y diseñar estrategias de prevención en zonas concretas, incluso en épocas determinadas del año.
Este proyecto ha sido financiado por CEVA Salud Animal (España) y la Fundación General de la Universidad de Salamanca.
Bibliografía:
- Escobar, L. E. (2020). Ecological niche modeling: an introduction for veterinarians and epidemiologists. Frontiers in Veterinary Science, 7, 519059.
- Esteves-Guimarães, J., Matos, J. I., Leal-Sousa, B., Oliveira, P., Lobo, L., Silvestre-Ferreira, A. C., Soares, C. S., Rodríguez-Escolar, I., Carretón, E., Morchón, R., Fontes-Sousa, A. P., Montoya-Alonso, J. A. (2024). Current state of canine heartworm in Portugal. Animals, 14(9), 1300.
- González-Mohino, E. I., Rodríguez-Escolar, I., Balmori-de la Puente, A., Collado-Cuadrado, M., Carretón, E., Montoya-Alonso, J. A., Morchón, R. (2025). Monthly analysis of the current risk of heartworm transmission in Portugal and Spain through ecological niche modeling as a control measure. Current Research in Parasitology & Vector-borne Diseases, 8, 100330.
- Montoya-Alonso, J. A., Morchón, R., García-Rodríguez, S. N., Falcón-Cordón, Y., Costa-Rodríguez, N., Matos, J. I., Rodríguez-Escolar, I., Carretón, E. (2022). Expansion of canine heartworm in Spain. Animals, 12(10), 1268.
- Morchón, R., Rodríguez-Escolar, I., Lambraño, R. E. H., Agudo, J. Á. S., Montoya-Alonso, J. A., Serafín-Pérez, I., Fernández-Serafín, C., Carretón, E. (2023). Assessment Heartworm Disease in the Canary Islands (Spain): Risk of Transmission in a Hyperendemic Area by Ecological Niche Modeling and Its Future Projection. Animals, 13(20), 3251.
- Rodríguez-Escolar, I., Balmori-de la Puente, A., Infante González-Mohino, E., Collado-Cuadrado, M., Carretón, E., Montoya-Alonso, J. A., Morchón, R. (2025). Assessment of the monthly risk of dirofilariosis infection in Europe and its projection to 2100 under climate change from a One Health perspective. Parasites & Vectors, 18, 516.





